

Les Amphibiens
Dans la nature actuelle, les Amphibiens sont représentés par trois groupes: les Anoures (grenouilles et crapauds), les Urodèles (salamandres et tritons) et les Gymnophiones (apodes). Ces trois groupes descendent très vraisemblablement d’un ancêtre commun qui leur est propre, ce qui fait des Amphibiens actuels un groupe monophylétique, souvent aussi désigné sous le nom de Lissamphibiens. Malgré leur grande variété de formes, les Amphibiens actuels, ou Lissamphibiens, partagent plusieurs caractères spécialisés que l’on ne retrouve chez aucun autre groupe de Vertébrés et qui témoignent de leur ascendance commune: les dents pédicellées et bicuspides, le mode d’accommodation visuelle, les glandes pluricellulaires de la peau (séreuses et muqueuses) et quelques spécialisations de l’oreille interne.
Par extension, on a placé dans les Amphibiens tout un ensemble hétérogène de Tétrapodes primitifs fossiles qui n’ont en commun avec les Amphibiens actuels que des caractères généraux de Tétrapodes, et dont certains sont plus primitifs que les Lissamphibiens, tandis que d’autres sont plus proches des Amniotes. De ce fait, le terme Amphibien, lorsque ces fossiles sont pris en compte, ne désigne plus un groupe monophylétique, ou clade, mais un groupe paraphylétique, ou grade, intermédiaire entre Poissons et Amniotes.
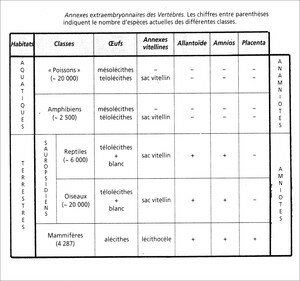
De fait, sauf de rares exceptions, les Amphibiens pondent dans l’eau douce et c’est là que leur développement embryonnaire se fait. Les larves sont dépourvues d’annexe embryonnaire et les Amphibiens sont anamniotes.
Ainsi, que ce soit au cours de leur phylogenèse ou au cours de leur ontogenèse, ces animaux se partagent entre une existence aquatique et une existence terrestre, justifiant pleinement leur nom d’Amphibiens.
1. Structures anatomiques et fonctions
Peau
La peau des Amphibiens actuels est molle, humide et nue, sauf chez plusieurs Gymnophiones (Caecilia d’Amérique du Sud par exemple) qui possèdent de petites écailles cycloïdes, imbriquées dans le derme. Les Stégocéphales avaient de grandes plaques osseuses sur la surface du crâne et sur le dos. Aujourd’hui, en dehors du bec corné des têtards, les phanères des Amphibiens se réduisent aux griffes – présentes à l’extrémité des doigts de certains Urodèles (Siren) et Anoures (Pipa, Pelobates) – ou à quelques formations cornées leur donnant un aspect parfois monstrueux (Ceratophrys, Megaphrys).
Souvent fortement pigmentés, les téguments possèdent de vives couleurs (triton marbré, salamandre, sonneur, etc.). Certaines glandes épidermiques sont muqueuses; répandu sur la peau de l’animal, le mucus la maintient humide en permanence: il joue un rôle lors du rapprochement des sexes ou pendant la locomotion (cas des rainettes arboricoles ). Des glandes séreuses (glandes parotoïdes des Salamandridés ou des Bufonidés) produisent un venin, liquide laiteux contenant des alcaloïdes toxiques; par suite de l’absence des dispositifs anatomiques nécessaires, il n’est ni projeté ni inoculé et constitue seulement une protection contre les prédateurs. Les glandes peuvent enfin être mixtes.
Après la métamorphose, les Amphibiens subissent périodiquement des mues. L’exuviation affecte la couche épidermique sur le corps entier de l’animal. Elle ne semble pas liée à une augmentation de taille et dépend du fonctionnement de la glande thyroïde.
Squelette
Les os sont fibreux (faisceaux de grosses fibres parallèles) constituant un squelette relativement léger qui peut supporter cependant des tractions ou des poussées très fortes, en particulier chez les Anoures, lors du saut.
Crâne
Il est caractérisé chez les formes actuelles par une importante fenestration qui affecte aussi bien le toit que le palais. En s’articulant avec la région postérieure du crâne, la première vertèbre cervicale constitue un atlas. Phylogénétiquement, c’est une innovation dans la série des Vertébrés, car la tête possède ainsi une certaine mobilité par rapport au tronc: on peut voir là l’ébauche d’un cou. Une autre importante nouveauté est l’apparition, pour la première fois, de l’oreille moyenne. L’accolement d’une évagination pharyngienne avec l’épiblaste entraîne la formation du tympan. Un os du deuxième arc viscéral (l’hyomandibulaire) ne participe plus à l’articulation de la mandibule comme chez les Poissons, mais vient se placer, au cours de l’ontogenèse, entre la fenêtre ovale de la capsule otique et la membrane tympanique: c’est la columelle. Cet ensemble anatomique assure la transmission des vibrations sonores en milieu aérien
Colonne vertébrale
Comme chez les autres Vertébrés, le squelette axial s’organise autour de la corde dorsale, par ossification dermique. L’adaptation à la vie terrestre correspond à un renforcement de la colonne vertébrale. La vertèbre typique des Amphibiens actuels a ses faces crâniale et caudale concaves: elle est dite pour cette raison amphicœle. L’interprétation de sa structure nécessite la connaissance de la vertèbre des Crossoptérygiens. De cette vertèbre de type rachitome dérive, selon les paléontologistes, la vertèbre stéréospondyle
L’arc neural, dorsal, est complètement ossifié et soudé au corps vertébral comme chez tous les Tétrapodes. L’arc hémal, ventral, qui n’existe que dans la région caudale, figure un vestige ichthyen.
Les vertèbres s’articulent entre elles grâce aux deux zygapophyses placées à la base de l’arc neural. Deux autres apophyses situées sur le corps s’articulent avec les côtes (bien développées seulement chez les Urodèles). Cependant, même lorsque les côtes existent, il n’y a jamais de véritable cage thoracique: elles ne se soudent pas au sternum.
Membres
Le passage de la vie aquatique à la vie terrestre est anatomiquement associé à la réalisation du membre chiridien (ou chiridium). Ce membre (antérieur ou postérieur), construit sur le même plan chez tous les Tétrapodes, se compose de deux unités principales: l’archépodium proximal (stylopode + zeugopode), dont on retrouve les homologues dans le membre des Crossoptérygiens, et le néopodium distal (basipode + métapode + acropode), qui apparaît chez les Tétrapodes. En plus de son rôle locomoteur, le membre chiridien soulève le corps. Sa position est transversale chez les Amphibiens.
Les formes actuelles possèdent un humérus plus léger que celui des formes ancestrales, mais l’articulation stylo-zeugopodiale a conservé une structure primitive. Le radius et l’ulna (ou cubitus) ont à peu près la même importance. Distalement, étant donné la direction de l’autopode, ces deux os sont obligés de venir côte à côte, ce qui amène l’ulna à se placer derrière le radius: c’est la pronation. La «main», au carpe réduit, n’a que quatre doigts chez les formes actuelles comme chez certaines formes fossiles. Il est plus probable que cet état est secondaire, car les plus anciens Tétrapodes connus possèdent de six (Ichthyostega, Tulerpeton) à huit doigts (Acanthostega).
Le membre postérieur montre les mêmes particularités que le membre antérieur. Ayant moins d’efforts à supporter (sauf en cas d’adaptation spéciale), sa construction est plus légère.
L’articulation de la main, ou du pied, ne se fait pas entre le zeugopode et le basipode comme chez les Vertébrés supérieurs, mais entre basipode et métapode.
Ceintures
La ceinture pectorale ou scapulaire, qui est libre, comme chez les Mammifères. Elle se compose principalement d’un scapulo-coracoïde
La tête du fémur s’emboîte dans la cavité acétabulaire (ou cotyloïde) de la ceinture pelvienne, soudée à la colonne vertébrale. Le bassin des Urodèles reste cartilagineux (Protée) ou partiellement ossifié (Tylotriton). L’adaptation au saut s’accompagne chez les Anoures d’une profonde modification de la morphologie du bassin. Il n’y a ni membres ni ceintures chez les Gymnophiones (Apodes).
Musculature
La musculature du tronc, que l’on qualifie de pariétale, est moins importante que chez les Poissons. Elle perd en grande partie le rôle qu’elle jouait lors des mouvements ondulatoires du corps. Mais, en même temps, elle acquiert de nouvelles fonctions: maintien de la rigidité de l’axe vertébral, réalisation des flexions latérales et dorso-ventrales du corps. La disparition quasi totale du septum horizontal coïncide avec une forte réduction de la musculature troncale épaxiale à l’avantage de la portion hypaxiale. La première perd pratiquement tous ses rapports avec les myoseptes et les vertèbres, tandis que la seconde se développe avec l’apparition du membre chiridien.
La musculature des membres, ou appendiculaire, n’a plus rien de commun avec celle des Poissons, les modes de locomotion étant tout à fait différents. C’est d’ailleurs pourquoi la musculature appendiculaire des Anoures n’est pas identique à celle des Urodèles. Ainsi, les extenseurs sont plus nombreux et plus volumineux chez les premiers. Cependant, la musculature du membre postérieur est, chez les formes primitives, à peu près la même. Ce fait est un argument pour les paléontologistes qui défendent l’existence d’un ancêtre commun aux deux ordres. De façon générale, la musculature appendiculaire des Amphibiens se compose de muscles courts, profonds, et de muscles longs, superficiels. Dans le cas particulier de la main et du pied, l’ontogenèse comme la phylogenèse montrent un remplacement progressif des muscles courts par des muscles longs.
Bouche
Elle sert autant à conduire l’air vers l’appareil respiratoire que les aliments vers le tube digestif. En effet, les quelques glandes intermaxillaires qui y débouchent ne sécrètent pratiquement pas d’enzymes digestifs, et les faibles dents qui la garnissent peuvent tout juste jouer un rôle préhenseur. C’est donc une simple ouverture, destinée à la capture des proies qui viennent s’engluer sur le mucus abondant qui couvre la langue. Dans le plafond buccal antérieur s’ouvrent les choanes (narines internes). La denture est très faible chez tous les Amphibiens. Chez certaines formes fossiles, la dentine présente des circonvolutions et des replis qui comblent la cavité pulpaire: c’est la structure de type «labyrinthodonte», héritée des ancêtres ostéolépiformes des Tétrapodes.
Tube digestif
Il présente quelques caractères particuliers. L’œsophage a un épithélium pseudo-stratifié à cellules muqueuses et à cellules ciliées. L’épithélium stomacal est simple. En dehors des cellules épithéliales muqueuses, chaque glande gastrique est uniquement composée, comme chez les Poissons, d’un seul type de cellules séreuses sécrétant à la fois la pepsine et l’acide chlorhydrique. L’épithélium intestinal, le foie et le pancréas présentent des caractères histologiques qu’on retrouve chez tous les Vertébrés.
Les Amphibiens adultes se nourrissent normalement d’insectes. Une grande partie des aliments est mise en réserve sous forme de graisse dans des endroits très divers, et sous forme de glycogène dans le foie. Ces réserves sont utilisées pendant la longue hibernation qui caractérise le cycle biologique des Amphibiens.
Sang
Les éléments figurés du sang, formés dans la rate, le foie, la moelle rouge, se répartissent en trois catégories: les leucocytes, les thrombocytes (ou plaquettes) et les érythrocytes. Ces derniers sont ovales et nucléés; leur diamètre est assez important puisqu’il peut atteindre jusqu’à 87 mm chez Amphiuma. La glycémie est de l’ordre de 0,5 gramme par litre. Le pH sanguin n’est pas stable, la réserve alcaline étant basse. L’homéostasie est donc encore fort imparfaite: aussi la calcémie est-elle deux fois plus importante en été qu’en hiver. Cela met en relief l’influence du milieu sur les régulations physiologiques – notion capitale chez les Amphibiens.
Appareil respiratoire
Chez les Amphibiens adultes, il est de type pulmonaire. Mais la présence d’une paire de poumons chez ces animaux a une importance qui est plutôt d’ordre anatomique et phylogénétique que d’ordre fonctionnel. En effet, les larves possèdent toutes une respiration branchiale et certains Urodèles conservent des branchies toute la vie. De plus, chez l’adulte, la peau a un rôle respiratoire parfois prédominant.
L’appareil pulmonaire se compose d’abord de la glotte, qui s’ouvre dans l’arrière-bouche. Elle se prolonge par une courte trachée, sauf chez les Anoures. Le larynx existe chez tous les Amphibiens; dans les cas les plus simples, il se compose d’une paire de cartilages (aryténoïde et cricoïde). Chez les Anoures, il prend une importance toute particulière dans l’émission des sons.
Chaque poumon (qui embryologiquement dérive du 7e diverticule pharyngien du têtard) est un sac à paroi mince. De celle-ci partent des cloisons de premier ordre qui divisent la cavité centrale en plusieurs grandes chambres. Des cloisons de deuxième ordre, partant des premières, découpent ces chambres en alvéoles. L’épithélium respiratoire est simple, cubique: les cellules laissent entre elles des espaces pour les capillaires sanguins qui sont ainsi directement en contact avec l’air des alvéoles.
On distinguait, classiquement, trois types de respiration chez les Amphibiens: cutanée, bucco-pharyngée et pulmonaire. La respiration bucco-pharyngée se traduirait extérieurement par l’espèce de «déglutition», très facile à observer sur une grenouille: les échanges gazeux se feraient au niveau de l’épithélium bucco-pharyngé richement vascularisé. On considère que ce phénomène est d’ordre olfactif et agit comme une pompe foulante pour remplir d’air les poumons. Dans la respiration pulmonaire (fonctionnellement associée à la précédente), l’air est aspiré dans la cavité buccale par les narines, puis chassé dans les poumons après fermeture des narines et élévation du plancher buccal; enfin, il est expiré lorsque les poumons reviennent sur eux-mêmes. La respiration cutanée est un fait bien établi chez tous les Amphibiens. Leur peau molle, humide et fortement vascularisée, réalise une membrane très perméable aux gaz. Aussi, les Amphibiens doivent-ils constamment rester dans un milieu dont le degré hygrométrique est élevé.
Chez les espèces dont les poumons sont réduits (Cryptobranchus) ou absents (Spelerpes), des plis ou des diverticules cutanés, abondamment vascularisés, apparaissent en plusieurs endroits du corps.
Appareil circulatoire
Du point de vue phylogénétique, l’apparition de la respiration pulmonaire a coïncidé avec des modifications importantes. Par rapport au cœur des Poissons, la forme en «S» s’accentue, reportant l’atrium à l’avant du ventricule, et le sinus veineux en avant de l’atrium. Celui-ci est subdivisé en deux cavités, les «oreillettes», l’une droite, l’autre gauche, par un septum interauriculaire, incomplet chez les Urodèles
Les six arcs aortiques primitifs sont remaniés au cours du développement embryonnaire. Les deux premiers disparaissent toujours, mais le troisième constitue l’arc carotidien qui irrigue la tête, et le quatrième forme l’arc systémique, dont les deux crosses initiales se réunissent dorsalement pour former l’aorte; le cinquième n’existe que chez les Urodèles; le sixième, l’arc pulmo-cutané, se divise en artère cutanée et en artère pulmonaire conduisant le sang réduit du ventricule aux poumons.
Le sang noir venant de la tête est drainé par les veines jugulaires, puis les veines caves antérieures
Il convient de remarquer que l’appareil circulatoire des Urodèles rappelle davantage celui des Dipneustes que celui des Anoures. Cet argument a longtemps été avancé par les paléontologistes partisans de la filiation: Dipneustes-Urodèles.
Le système lymphatique des Amphibiens se compose de vastes sinus sous-cutanés bien individualisés. La lymphe qui y circule est mue par de petits organes pulsatiles placés sur le trajet des vaisseaux; ce sont les cœurs lymphatiques (on en compte jusqu’à 200). Systèmes sanguin et lymphatique sont en communication étroite. Il en résulte un renouvellement constant du plasma en mouvement.
Organes sensoriels
Peut-être plus que tous les autres Vertébrés, les Amphibiens possèdent une grande variété d’organes sensoriels. La peau a conservé en général un caractère ichthyen, la ligne latérale, qui intervient dans les réflexes d’équilibration et de posture. Elle est également sensible, grâce à des récepteurs spécialisés, au stimulus d’origine mécanique, chimique, lumineuse, ainsi qu’à la chaleur, au froid et à la douleur.
Les organes du goût sont localisés au palais et à la langue. Ce sont de petits amas (bourgeons) de cellules cylindriques, innervés par des filets de nerfs crâniens V, VII, IX et X. Il n’y a pas, chez les Amphibiens, de nerf gustatif individualisé.
Chaque organe olfactif comprend trois chambres reliées les unes aux autres, les deux chambres extrêmes débouchant d’une part à l’extérieur, par la narine, d’autre part dans la cavité buccale, par le choane. Ce dernier est entouré par une différenciation spéciale de l’épithélium olfactif, qui constitue l’organe voméronasal (ou organe de Jacobson).
L’œil est de taille très variable chez les Amphibiens (il suffit en effet de comparer une grenouille et un triton). Certains, comme le protée, sont même aveugles. La protection de l’œil est assurée par la présence de paupières et de glandes lacrymales. L’accommodation ne se fait pas par un changement de forme du cristallin, mais par un changement de position du globe oculaire dans l’orbite, nécessitant l’existence de muscles spécialisés. La rétine présente une structure histologique caractéristique de l’ensemble des Vertébrés. En particulier, ses différents types synaptiques lui confèrent un intérêt tout particulier.
L’oreille interne ne présente, par rapport à celle des Chondrichthyens, qu’une lagena un peu plus développée par suite de l’existence de deux nouvelles papilles: la papilla basilaris et la papilla amphibiorum (spécifique aux Amphibiens). L’espace périlymphatique n’entoure que le vestibule et les canaux semi-circulaires. L’oreille moyenne est constituée par la columelle, peu ossifiée
Système nerveux
Dans ses grandes lignes, l’architectonie du névraxe des Amphibiens est suffisamment simple pour être considérée comme primitive.
Moelle: la substance grise médullaire est déjà organisée en cornes dorsales et cornes ventrales contenant les grands neurones moteurs. La substance blanche est constituée principalement par les voies des sensibilités protopathique et épicritique (celle-ci n’existait pas chez les Poissons), et par les voies motrices descendantes, limitées, ici, au système extra-pyramidal.
Bulbe rachidien: la région ventrale, motrice, contient les noyaux des nerfs crâniens III à XII (ce dernier apparaît chez les Amphibiens en relation avec l’existence d’une langue musculeuse). L’aire sensitive dorsale reçoit, d’une part, les filets des nerfs crâniens V, VII, IX et X, et envoie, d’autre part, un ensemble de fibres (lemnisque latéral) au toit du mésencéphale
Cervelet: il est réduit à une lame mésiale (corps cérébelleux), centre d’intégration des réflexes de posture et d’équilibre, flanquée de deux auricules (archicerebellum) qui appartiennent physiologiquement au système acoustico-latéral.
Mésencéphale: il représente un centre d’intégration important du cerveau des Amphibiens, recevant des influx acoustico-latéraux, optiques, proprio- et extéroceptifs, et même, olfactifs.
Diencéphale: il est surmonté, dorsalement, par l’épiphyse (organe pinéal), qui est reliée à l’organe frontal. Celui-ci est peut-être par sa situation et son origine l’homologue de l’œil pinéal reptilien. Ventralement, l’hypothalamus constitue un centre de corrélation entre les influx gustatifs et olfactifs, relié à l’hypophyse, dont le lobe nerveux commence à ébaucher une tige pituitaire et une pars tuberalis; le thalamus, important centre optique (corps genouillés latéraux), se place en position intermédiaire.
Hémisphères cérébraux: totalement démunis de cortex, ils sont constitués chacun par deux zones cellulaires; dorsalement le pallium, et ventralement le subpallium, qui est essentiellement un centre olfactif. Ce dernier envoie ses influx au diencéphale par les importants cordons télencéphaliques basaux dont les fibres se croisent dans la commissure antérieure. Du pallium partent des fibres assimilables à celles du fornix mammalien. Chaque hémisphère reçoit, en plus des informations olfactives, des afférences visuelles et peut-être acoustiques.
Système nerveux autonome: il ne présente guère de différence avec celui des Mammifères. Il faut cependant noter qu’il existe peu de ganglions prévertébraux orthosympathiques. On admet, généralement, que le mécanisme émotionnel neurovégétatif, bien que difficile à constater chez ces animaux, fait intervenir les mêmes centres que chez les Vertébrés supérieurs.
Glandes endocrines
Seules sont indiquées ici les glandes non étudiées par ailleurs.
Adénohypophyse: elle sécrète d’une part des stimulines et d’autre part l’intermédine, qui intervient dans les variations de pigmentation.
Parathyroïdes : dans la série des Vertébrés, elles apparaissent chez les Amphibiens où elles interviennent dans le métabolisme du calcium et du phosphore.
Glande adrénale: constituée par un ensemble de petits corpuscules placés en avant des reins, elle se compose de trois types de cellules dont la spécialisation est loin d’atteindre celle de la glande surrénale des Mammifères; néanmoins, on reconnaît des cellules corticales, des cellules (de Stilling) particulières aux Amphibiens, et des cellules chromaffines responsables de la sécrétion d’adrénaline dont le rôle cardio-accélérateur a été précisément mis en évidence chez les Amphibiens (expériences de Loewi).
Toutes les glandes endocrines ont une activité cyclique saisonnière.
Appareil excréteur
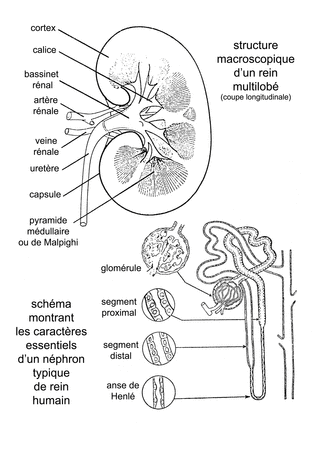
Les reins sont situés très dorsalement, de part et d’autre de la colonne vertébrale. Ce sont des reins secondaires (mésonéphros). Au cours du développement, leur partie antérieure dégénère chez la femelle, alors qu’elle est annexée par l’appareil génital chez le mâle. La partie postérieure possède, seule, la fonction excrétrice.
La diurèse est très importante et dépend de la température. L’urine, très diluée, présente une pression osmotique toujours inférieure à celle du sang. La perte en eau est considérable et l’excrétion des Amphibiens paraît, de façon générale, mal adaptée à la vie terrestre.
Appareil génital
Chez le mâle, le testicule est un agrégat d’ampoules contenant les cellules germinatives et les cellules de Sertoli. Entre ces ampoules, un tissu interstitiel sécrète l’hormone androgène responsable de l’apparition des caractères sexuels secondaires, parfois très marqués (triton).
L’appareil génital mâle
L’appareil génital femelle
Les ovules sont expulsés dans la cavité générale du tronc puis repris par le pavillon cilié placé à l’extrémité crâniale de l’oviducte ou canal de Muller.
Les voies urinaires et génitales s’ouvrent, ainsi que le rectum, dans une chambre ventrale que l’on appelle le cloaque.
La ponte et la fécondation présentent des caractères extrêmement variables selon les ordres, ou même les familles.
Développement
De nombreuses expériences d’hybridation ont été réalisées chez les Amphibiens, qui constituent toujours un matériel favorable à toute expérimentation. En outre, il faut noter l’existence d’hybrides naturels qui soulève des problèmes de classification, par exemple chez les tritons. Après être sorti de l’œuf, au stade «bourgeon caudal» en général, l’embryon devient une larve, bien connue chez la grenouille sous le nom de têtard. C’est dans l’eau que se déroule sa vie, qui comporte une succession de transformations anatomiques et physiologiques dont l’ensemble constitue la métamorphose. Les particularités de la reproduction et du développement seront rattachées à l’étude des ordres.
Métabolisme
Dans leurs grandes lignes, les mécanismes qui interviennent (synthèses, dégradations, utilisation de l’énergie, métabolisme intermédiaire) sont identiques à ceux des autres Vertébrés.
La poïkilothermie (température variable), qui présente par ailleurs des inconvénients, permet cependant aux Amphibiens de résister à de longs jeûnes, aux faibles pressions en oxygène (on trouve des Anoures à plus de 5 000 m dans l’Himalaya) et à la perte d’énergie par rayonnement de chaleur.
L’étude du métabolisme de ces animaux (qui prend de plus en plus d’importance) ne peut que contribuer, par suite de leur position phylogénétique, à mieux faire comprendre celui des êtres homéothermes.
2. Ecologie
On peut envisager l’écologie des Amphibiens sous deux aspects différents. Du point de vue biologique, ces animaux présentent un cycle sexuel annuel qui se superpose à un comportement également cyclique. L’hiver, ils sont au repos sexuel et entrent en état d’hibernation; ils se dissimulent, s’enterrent même, dans les berges des mares ou des ruisseaux, sous les roches ou sous les écorces d’arbres. Au printemps, l’activité sexuelle reprenant, les animaux sortent de leur retraite.
On pourrait aussi classer les Amphibiens en trois catégories, selon leur habitat. Certaines espèces toujours aquatiques (comme la grande salamandre du Japon ou l’Ambystoma) passent leur vie dans les eaux courantes. D’autres, toujours terrestres, comme la salamandre noire d’Europe ou le Nectophrynoïdes d’Afrique, sont ovovivipares: la vie larvaire est «intra-utérine», gardant ainsi son caractère essentiellement aquatique ; à l’éclosion, les jeunes naissent déjà adaptés à la vie terrestre. Cependant les espèces les plus nombreuses sont véritablement amphibies (grenouille, crapaud, rainette, triton) et ne s’éloignent jamais des mares ou des ruisseaux où elles reviennent obligatoirement pour la fécondation
Parasites et maladies. De nombreux parasites, de toutes tailles, infestent certains organes. Ainsi, la peau est attaquée par des champignons provoquant des mycoses, souvent mortelles. La maladie dite «des pattes rouges» (congestion des vaisseaux sanguins superficiels de la face ventrale des pattes) est d’origine bactérienne.
Parmi les Flagellés, on trouve des Eugléniens dans le tube digestif et des Trypanosomidés dans le sang. Les Opalinidés (Ciliés) sont communs dans le tube digestif, avec des Amibiens et des Sporozoaires. Ces derniers, très répandus chez les Amphibiens, envahissent de nombreux organes en particulier les reins.
Les Helminthes sont représentés, entre autres, par l’Ascaris, certains Acanthocéphales, le Tوnia, tous localisés au tube digestif. Le plus commun est un Polystonum (Trématode) qui abonde dans la vessie des grenouilles.
Ennemis. Dès l’œuf, les Amphibiens sont les victimes d’un nombre considérable de prédateurs. Le poisson-chat, par exemple, se nourrit de leurs œufs et de leurs têtards. Ceux-ci sont également la proie d’insectes aquatiques carnivores (Dytique). Adultes, ils sont la nourriture favorite des serpents. Enfin, les hérons, les rats, les chats, l’homme contribuent à leur destruction.
3. Evolution
On considère désormais que, des trois groupes actuels d’Amphibiens, ce sont les Urodèles et les Anoures qui sont les plus étroitement apparentés et sont regroupés sous le nom de Paratoïdes, tandis que les Gymnophiones constituent leur groupe frère
Les affinités des Lissamphibiens avec les autres groupes de Tétrapodes primitifs fossiles sont encore obscures et sont l’objet de nombreuses controverses. Gardiner (1983) a remis à l’honneur l’idée d’une relation étroite entre Lissamphibiens et Lépospondyles, un petit groupe d’Amphibiens paléozoïques comprenant des formes à allure de tritons (Nectridiens) et parfois serpentiformes (Aistopodes). Cependant, beaucoup de paléontologues soutiennent encore l’hypothèse selon laquelle les Paratoïdes (Anoures + Urodèles) descendraient d’un petit groupe de Temnospondyles permiens, les Dissorophidés, tandis que les Gymnophiones seraient issus d’un autre groupe fossile, les Microsaures (Rage, 1986; Milner et al., 1986). Gardiner, en revanche, récuse une relation étroite entre Lissamphibiens et Temnospondyles, et considère l’ensemble Lissamphibiens + Lépospondyles comme plus étroitement apparenté aux Amniotes qu’aux autres Amphibiens fossiles.
Quant aux autres Amphibiens fossiles, les « Stégocéphales » ou « Labyrinthodontes », ils se subdivisent dès le début du Carbonifère (et, peut-être déjà, dès le Dévonien) en deux grands ensembles: d’une part les Anthracosaures, aux vertèbres de type embolomère et d’où sont issus les Amniotes, et d’autre part les Temnospondyles, qui peuplèrent les zones humides du globe jusqu’au début du Jurassique et dans lesquels s’enracineraient peut-être les Lissamphibiens. En dehors de ces deux grands ensembles, trois petits groupes, les Ichthyostégides, les Loxommatides et les Crassigyrinides, représentent les plus primitifs des Tétrapodes connus.
Le passage du grade amphibien au clade amniote est marqué essentiellement par l’apparition de l’œuf amniotique, événement qu’il est difficile de dater, sauf, peut-être, grâce à l’horloge moléculaire. La paléontologie, quant à elle, permet d’affirmer qu’il existait déjà des reptiles – donc des Amniotes – au Carbonifère moyen. Pour Gardiner, tous les Anthracosaures et les Microsaures seraient déjà à placer parmi les Amniotes, ce qui repousse l’origine des Amniotes à la fin du Dévonien. Au sein des Anthracosaures, ce sont les Seymouriamorphes qui partagent avec les Amniotes typiques le plus grand nombre de spécialisations (structure des vertèbres, du crâne) et semblent déjà montrer, par la réduction de leurs sillons sensoriels de la ligne latérale, un certain affranchissement vis-à-vis du milieu aquatique.
Tous ces grands Amphibiens fossiles à allure de crocodiles qui peuplaient les marécages, voire les lagunes marines au Paléozoïque, se sont éteints à la fin du Trias ou au tout début du Jurassique.
























/https%3A%2F%2Fprofilepics.canalblog.com%2Fprofilepics%2F2%2F2%2F220176.jpg)