

Ontogenèse, Morphogenèse
L’immense majorité des espèces vivantes qui peuplent actuellement la biosphère est constituée d’organismes pluricellulaires diploïdes et doués de sexualité. Il paraît donc raisonnable d’admettre que le fait d’être pluricellulaire, diploïde et sexué confère aux organismes des avantages réels.
En nous plaçant dans une perspective évolutionniste, une population d’organismes a d’autant de chance de s’adapter aux variations du milieu où elle vit que ses possibilités de variations sont plus grandes.
1. Propriétés majeures des organismes pluricellulaires
L’observation montre que la pluricellularité s’accompagne d’un phénomène de spécialisation cellulaire. En effet, la plupart des plantes et des animaux sont constitués de cellules spécialisées dans diverses fonctions : soutien, revêtement, perception, locomotion, coordination, etc.…Les plus complexes des pluricellulaires, comme les Mammifères, produisent chacun un peu plus de 200 types de cellules différents.
Les organismes pluricellulaires présentent une organisation supracellulaire spécifique : les cellules parfois très nombreuses (1012 dans notre espèce) sont agencées suivant un plan strictement spécifique. L’organisation d’une plante ou d’un animal s’établit progressivement au cours de la période de formation d’un individu nouveau à partir d’un œuf, d’une spore ou d’un bourgeon.
La rigueur de cette organisation peut être mise en évidence par l’étude expérimentale d’animaux doués de régénération. L’expérience montre que la régénération d’un organisme comprend fondamentalement trois étapes :
1. L’agrégation au niveau de la blessure de cellules indifférenciées.
2. La croissance par divisions mitotiques de cet agrégat (blastème).
3. La différenciation et l’ordonnance de ce blastème.
En règle générale, l’individu pluricellulaire complet et fonctionnel est construit à partir d’une masse de cellules indifférenciées produite par la division d’un œuf, la formation d’un bourgeon ou celle d’un blastème de régénération.
L’ensemble des phénomènes conduisant à la différenciation ordonnée d’un germe indifférencié est désigné par le terme de morphogenèse.
2. Modes de formation des organismes pluricellulaires
A priori, on peut imaginer deux modes d’associations de cellules conduisant à la formation d’un individu pluricellulaire nouveau :
· Les Acrasiales, dont leur biologie les situe aux frontières des Protozoaires Amibiens et des pluricellulaires. L’espèce Dictyostelium, se reproduit à l’aide de spores disséminées dans le milieu naturel, de chaque spore émerge une amibe solitaire. Les amibes des Acrasiales ont la capacité de s’associer en un agrégat cohérent. Cette capacité est liée à l’émission d’un messager chimique, l’AMP cyclique. Les amibes agrégées se différencient. Leur différenciation dépend d’un ensemble de gènes. Ces gènes ne s’expriment que lorsque les amibes sont en contact et ils s’expriment différemment selon la position de chaque amibe. La destinée de chaque amibe dépend donc à la fois des propriétés de ses gènes et des propriétés du milieu qui l’environne.
· Plusieurs espèces de Spongiaires se reproduisent de manière asexuée en produisant des gemmules.
b. Association des cellules issues d’une cellule fondatrice initiale
Les mitoses successives d’une cellule initiale fondatrice (spore ou zygote) produisent des organismes pluricellulaires résultant de processus morphogènes.
Chez les Plantes terrestres et chez les Animaux, la cellule fondatrice est généralement l’œuf fécondé dont le développement présente de remarquables similitudes chez toutes les espèces. Le zygote commence par se transformer en un embryon pluricellulaire massif dont la croissance, l’organisation et la différenciation produiront finalement un individu adulte. L’étude de ces processus est le propre de l’embryologie.
.II. Embryogenèse d’une espèce animale type
1. Segmentation
La segmentation est un processus qui transforme l’œuf fécondé en une masse de cellules embryonnaires, les blastomères, dont le volume total est à peu près égal à celui de l’œuf initial. Les divisions de segmentation sont des mitoses singulières, à la fois rapides et rapprochées, à interphases très courtes.

32. Gastrulation
La gastrulation est la mise en place dans l’embryon des différentes strates embryonnaires fondamentales, les feuillets, dont vont dériver les organes de l’animal. Elle se manifeste par des mouvements des territoires embryonnaires tandis que les mitoses, normalisées se poursuivent. L’embryon qui est le siège de ces mouvements est une gastrula.

3. Organogenèse
Au cours de l’organogenèse, les feuillets vont se morceler en territoires plus restreints, ébauches des futurs organes. Les cellules qui constituent ces ébauches ont toujours un aspect embryonnaire, indifférencié.
La mise en place des ébauches neurales est la neurulation. L’embryon qui en est le siège est la neurula. La plaque neurale, d’origine ectodermique, se déprime en une gouttière allongée, élargie à son extrémité antérieure. Les bords de la gouttière se rapprochent peu à peu, se rejoignent et se soudent. Il se forme ainsi un tube neural.
* La métamérisation
Dans la jeune neurula, les lames mésodermiques gauche et droite s’étendent de manière continue d’un bout à l’autre de l’embryon. Ces lames vont se découper en une série de segments successifs : c’est la métamérisation.
4. Histogenèse
L’histogenèse est la transformation des ébauches embryonnaires en tissus ayant des structures et des fonctions spécifiques. Les blastomères se différencient en cellules spécialisées. Un tissu peut donc être défini comme un groupe de cellules ayant subi la même différenciation. Chez les Vertébrés, les cellules embryonnaires vont se différencier en plusieurs dizaines de types cellulaires différents. Si toutes les cellules d’un animal ont le même génome, la différenciation cellulaire implique que des gènes différents s’expriment dans chacun des types cellulaires.
5. Contribution des trois feuillets embryonnaires à l’édification des principaux organes des Vertébrés
Lorsque l’embryogenèse est achevée, les trois feuillets gastruléens auront finalement édifié les systèmes organiques suivants (voir tableau).
En général, les 5 grandes ébauches initiales de l’embryon produisent des familles de tissus qui leur sont caractéristiques :
· Épiderme et endoderme : épithéliums et glandes ;
· Neuroderme : tissus nerveux ;
· Cordoderme : cordocytes (grandes cellules vacuolisées) ;
· Mésoderme : tissus musculaires, cartilagineux, osseux, conjonctifs et sanguins.
Cette règle souffre cependant des exceptions. Les plus notables concernent des territoires épidermiques de la tête (microplacodes, crêtes neurales) qui peuvent neutraliser (et participer à la formation de ganglions et de nerfs crâniens) ou encore produire du tissu conjonctif et embryonnaire (ectomésenchyme).
ECTODERME MÉSODERME ENDODERME épiderme neuroderme cordoderme Mésoderme Sensu stricto Peau (phanères et glandes), ouvertures buccale et anale du TD. Adénohypophyse Organes de sens. Système nerveux. Neurhypophyse. Corde (organe larvaire chez les Vertébrés). Derme de la peau. Squelette. Musculature (volontaire et viscérale). Tissus de soutien des viscères. Mésentères. Appareil uro-génital (à l’exception des cellules germinales). Appareil circulatoire (vaisseaux, sang et cœur). Tube digestif et ses glandes annexes (glandes salivaires, foie, pancréas). Appareil respiratoire (branchies et poumons) et organes dérivés (thyroïde et parathyroïdes). Cellules germinales
.III. Notions d’embryologie comparée des animaux
1. Animaux di- et tridermiques
Chez les Cnidaires (environ 9000) espèces, il n’y a pas de mésoderme. La gastrule est un embryon didermique : l’endoderme interne est accolé à l’ectoderme périphérique. Dans les gastrules de tous les autres animaux, un mésoderme est interposé entre l’ecto- et l’endoderme. Du mésoderme dérivent presque tous les autres tissus interposés entre la peau et le tube digestif.
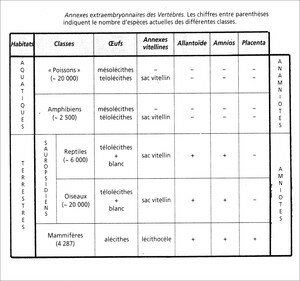
2. Annexes extra-embryonnaires des Vertébrés
Les annexes extra-embryonnaires se présentent sous l’aspect de poches et de diverticules, formés de feuillets émergeant de l’embryon mais en continuité avec ceux qui constituent le germe. Ce sont des dispositifs qui assurent la nutrition (sac vitellin, placenta), l’hydratation (amnios, allantoïde), la respiration (allantoïde, placenta) et l’excrétion de l’embryon (allantoïde, placenta).

3. Embryogenèse des Mammifères
En règle générale, les Mammifères sont vivipares et placentaires. Leurs œufs ou ovules, dépourvus de vitellus, sont microscopiques. Ces ovules sont fécondés dans le tiers supérieur de l’oviducte. La segmentation s’effectue pendant que l’œuf libre chemine de l’oviducte vers l’utérus. La morule libre devient creuse et se transforme en un blastocyste, constitué d’une couronne de blastomères, le trophectoderme, qui recouvre localement un bouton embryonnaire. L’implantation est l’étape de fixation du blastocyste par son trophectoderme à la paroi de l’utérus. Contrairement à ce qui se passe chez les Reptiles et les Oiseaux, l’édification des annexes extra-embryonnaires est très précoce : elle commence avant la gastrulation.
Chez les Mammifères, comme chez tous les Vertébrés, la paire d’ébauches des organes génitaux a la même structure chez tous les jeunes embryons, quel que soit leur sexe génétique : ces ébauches sont potentiellement bisexuées. La gonade embryonnaire contient les cellules germinales et comprend une région centrale, la médulla, et une région périphérique, le cortex. La médulla est susceptible d’évoluer en testicule et le cortex en ovaire. Les ébauches des gonades sont accompagnées de deux paires d’ébauches de conduits génitaux : les canaux de Mûller, ébauches des conduits femelles, et les canaux de Wolff, ébauches des conduits mâles. Le fœtus humain possède ces ébauches génitales, doubles et paires, à 7 semaines après la fécondation.
L’évolution ultérieure des ébauches génitales dépend du sexe génétique des individus. La présence d’un chromosome Y est l’élément déterminant dans le cas des Mammifères. En sa présence, la gonade fœtale se différencie en testicule. En son absence, elle produit un ovaire.
c. Anomalies sexuelles
Le sexe d’un individu adulte résulte donc d’un ensemble de phénomènes : génétiques, embryologiques, hormonaux, très complexes dont l’interaction fait apparaître dans les populations deux phénotypes sexuels, propres à perpétuer l’espèce. Mais des anomalies, des accidents peuvent se produire : nombre anormal d’hétérochromosomes, dérèglement hormonal lors de la période fœtale.
d. Chronologie de l’embryogenèse humaine
L’ovule est de 150 microns de diamètre. Le 5ème jour après la fécondation, la cavité de segmentation se creuse. Au 6ème jour, le blastocyste s’implante dans la paroi utérine. Pendant la 2ème semaine, tandis que le placenta se développe, le bouton embryonnaire se creuse de deux cavités superposées, l’amniotique et la vitelline. L’embryon gastrule pendant la 3ème semaine. A la fin de la 4ème semaine, la neurulation est achevée ; le diverticule allantoïdien s’est développé. L’organogenèse se poursuit jusqu’à la fin du 2ème mois. A 60 jours, la période embryonnaire est terminée. Pendant les 7 mois restants de la gestation, ou période fœtale, l’embryon va croître et se différencier.
.IV. Pouvoir de régulation et détermination embryonnaire
1. Pouvoir de régulation de l’œuf de triton
Si on sépare les 2 premiers blastomères -> 2 têtards normaux (taille réduite).
Si on met en contact 2 embryons (au stade 2 blastomères) -> 1 têtard unique (très grand).
La régulation est le pouvoir de réorganisation du germe (œuf ou embryon) qui a pour effet de produire un embryon normal après ablation ou ajout d’une de ses parties.
2. Détermination des régions embryonnaires (champs morphogènes)
La détermination embryonnaire correspond donc à une spécialisation d’une région embryonnaire qui s’exprime non par une différentiation morphologique mais par une capacité morphogène singulière pouvant être mise en évidence par l’expérimentation (ablations ou greffes). En d’autres termes, les cellules sont déterminées lorsque leur destin est fixé bien que leur aspect soit encore embryonnaire, non différencié. Plus grande est l’étendue des territoires déterminés d’un embryon, moins grand est son pouvoir de régulation.
3. Évolution du pouvoir de régulation au cours de l’embryogenèse
Les résultats des expériences d’ablation et de greffe ont plus de chance de réussite quand elles sont pratiquées sur des stades précoces du développement. Tout se passe donc comme si le pouvoir de régulation se perdait au cours du développement. Le pouvoir de régulation d’un embryon nous informe donc sur le degré de détermination de cet embryon. Lorsque ce pouvoir diminue, c’est que la détermination apparaît ; lorsqu’il est nul, c’est que la détermination est stable.
4. Généralité de la détermination embryonnaire chez les animaux
La plupart des espèces animales ont des œufs capables de régulation : Amphibiens, Oiseaux, Mammifères. Cependant, la détermination des champs morphogènes est plus ou moins précoce selon les espèces. Elle peut être totale, dès l’œuf indivis. Ainsi chez les Tuniciers, l’œuf est une mosaïque de territoires déterminés ; la détermination de tout le germe est liée à la fécondation.
Sa capacité de régulation est nulle. En somme, dans les œufs en mosaïque, la potentialité morphogène totale de chaque partie de l’œuf fécondé est égale à la potentialité exprimée au cours du développement embryonnaire normal. Dans les œufs à régulation, la potentialité totale est supérieure à la potentialité exprimée. Leurs différents territoires ne sont pas déterminés de manière synchrone. La régulation se manifestera d’autant plus longtemps que la détermination est tardive.
5. Caractères des territoires déterminés
Lorsqu’on centrifuge des œufs d’Amphibiens, la perturbation de la distribution normale des plastes est à l’origine de la formation d’embryons monstres (à 2 têtes). Aussi l’analyse cytologique des œufs à mosaïque, montre parfois une distribution inégale des mitochondries dans les différents territoires.
.V. Pouvoir d’induction et compétence des territoires embryonnaires
La détermination d’un champ embryonnaire peut se mettre en évidence en procédant soit à l’ablation de la région étudiée, soit à la greffe de cette région dans un autre embryon. En étudiant, dans cette perspective, le territoire présomptif cordal de jeunes gastrules de tritons, Spemann, mit en évidence un phénomène d’importance considérable en embryologie : l’induction.
1. Pouvoir inducteur du territoire cordal des Amphibiens
En greffant ce territoire dans l’ectoderme ventral d’une autre gastrule, il observa non seulement que le greffon se différenciait en corde, mais encore que le greffon cordal induisait la formation d’un embryon secondaire, aux dépens des tissus du porte-greffe. Le résultat de l’opération est l’obtention d’un embryon double. Aussi on a démontré que la greffe du cortex dorsal de l’œuf fécondé indivis peut induire la formation d’un embryon secondaire. Pour que l’induction puisse se produire, il faut que le territoire inducteur soit à proximité immédiate du territoire induit. L’induction est le processus selon lequel un territoire embryonnaire détermine l’évolution d’un autre territoire embryonnaire situé près de lui.
2. Compétence des territoires embryonnaires
Pour se manifester, l’induction exige donc la présence de deux territoires embryonnaires différents : l’un capable d’émettre des signaux inducteurs, l’autre capable de capter ces signaux et d’y répondre. La capacité de répondre à des signaux inducteurs est définie comme la compétence d’un territoire embryonnaire.
Les signaux inducteurs sont des substances chimiques de nature protéique dont certaines commencent à être identifiées grâce notamment aux méthodes performantes du génie génétique. La compétence des cellules embryonnaires est vraisemblablement liée à la présence dans la membrane plasmique de récepteurs spécifiques de ces substances inductrices.
Les résultats des expériences de culture de fragments conduisent à l’hypothèse que le territoire cordal émet plusieurs substances inductrices différentes. La qualité des récepteurs propres aux feuillets permet le captage de l’une ou l’autre de ces substances (ectoderme, mésoderme) ou au contraire leur surdité à leur égard (endoderme).
3. Caractères de l’induction primaire des Vertébrés
L’effet inducteur du territoire cordal sur l’ectoderme et le mésoderme s’observe chez tous les Cordés. C’est le phénomène d’induction primaire chez ces animaux. Le territoire cordal est le centre organisateur des embryons. Sa destruction produit la formation d’un embryon anormal, dépourvu de corde, de système nerveux, de tête, de muscles…
L’induction primaire présente un ensemble de propriétés communes à tous les Vertébrés :
L’induction primaire est régionalisée, le pouvoir inducteur n’est pas exactement le même sur toute son étendue,
Le pouvoir inducteur du territoire cordal diminue au fur et à mesure que ce territoire se différencie en corde.
Le pouvoir inducteur du cordoderme est transmissible.
L’induction primaire n’a pas de spécificité zoologique.
4. Nature chimique des substances inductrices des Vertébrés
L’existence de substances inductrices diffusibles a été postulée sur la base d’un ensemble de faits expérimentaux. Parmi ceux-ci citons :
le fait qu’un territoire inducteur tué demeure inducteur ;
le fait que les territoires inducteurs et compétents doivent être près l’un de l’autre pour qu’il y ait induction ;
le fait que le pouvoir inducteur est transmissible.
Par des techniques immunologiques, on a pu montrer que les substances inductrices sont très probablement des protéines. L’essor de la génétique moléculaire du développement a permis de confirmer ces conclusions par la découverte très récente d’espèces moléculaires diffusibles produites par l’organisateur au cours de la gastrulation.
5. Inductions en cascades et organogenèse
Dans certains cas, limités, la détermination et la différentiation d’un organe dépend uniquement des propriétés intrinsèques du territoire embryonnaire dont il est issu. Ces territoires sont doués d’auto-différentiation, comme l’endoderme qui évolue spontanément en structures intestinales.
Plus généralement, la genèse des organes se fait par une cascade d’inductions qui s’opèrent suivant une séquence bien définie au cours du temps, les territoires induits devenant à leur tour inducteurs. L’action inductrice de deux territoires est souvent réciproque. Enfin, la différenciation précise d’une ébauche d’organe résulte fréquemment d’inductions multiples.
La formation d’un œil de Vertébré illustre les phénomènes d’inductions secondaires, réciproques et multiples. L’œil se constitue à partir de territoires neuro-, méso- et épidermique de la région antérieure de l’embryon :
1. le mésoderme précordal induit le plancher de l’ampoule cérébrale : ce plancher émet 2 lobes, les vésicules optiques gauche et droite.
2. Au contact des vésicules optiques, l’épiderme céphalique s’épaissit de chaque côté de la tête en un disque, une placode, ébauche du cristallin.
3. Ces placodes agissent à leur tour sur les vésicules optiques qui se dépriment chacune en une cupule à double paroi. La paroi externe est l’ébauche de la couche pigmentée et la paroi interne, l’ébauche de la rétine.
4. Les cupules optiques induisent le mésoderme précordal à se différencier en sclérotique et en choroïde.
5. Enfin, l’effet conjugué de la rétine et du cristallin en formation induit l’épiderme sus-jacent à se différencier en cornée.
1. Culture, transfert et congélation d’embryons
Il existe trois méthodes pour obtenir des œufs en segmentation :
1. prélever sur l’animal des œufs fécondés et déjà divisés,
2. on féconde in vitro des œufs prélevés par ponction des follicules préovulatoires
3. prélever sur l’animal un petit fragment d’ovaire dont on extrait les ovocytes I qui sont fécondés après maturation in vitro.
Le prélèvement et le transfert d’embryons sont maintenant des pratiques courantes en médecine vétérinaire, notamment dans l’élevage des bovins. Elles permettent par exemple, de faire porter par des vaches de grande taille des embryons de la race « cul de poulain ». cette race est intéressante par la qualité des rôtis qu’elle fournit mais les femelles y sont de petite taille et vêlent difficilement de leurs veaux à l’arrière-train particulièrement rebondi.
Par ailleurs, les méthodes de congélation des œufs fécondés et des œufs segmentés sont maintenant bien au point. Elles permettent de différer dans le temps deux séquences d’une suite opératoire, par exemple la fécondation in vitro et le transfert d’embryon (dits FIVETE). C’est ainsi que lorsque la FIVETE est appliquée à une femme, il y a souvent production d’embryons surnuméraires qui peuvent être conservés par congélation (le plus souvent au stade 4 blastomères).
Régulation embryonnaire
Dans l’espèce humaine, l’existence de vrais jumeaux résultant du développement des deux moitiés d’un même embryon (jumeaux monozygotiques) est la preuve d’une régulation embryonnaire. Lorsque le partage se produit à l’éclosion, les deux hémiblastocystes s’implantent et se développent chacun en un conceptus complet. Les vrais jumeaux sont dans ce cas dichorioniques. Ils ont en effet des annexes extra-embryonnaires complètes et distinctes. Si le partage ne concerne que l’amas embryogène (cas le plus fréquent), les jumeaux vrais sont monochorioniques et diamniotiques. Ils partagent en effet le même placenta auquel ils sont rattachés par leur propre codon ombilical et disposent chacun d’un amnios. Enfin, si la séparation a lieu plus tard encore, au niveau du disque embryonnaire, les deux jumeaux partagent le même placenta et le même amnios (jumeaux monochorioniques, monoamniotiques).
Les faux jumeaux (jumeaux dizygotiques) résultent de la fécondation de deux ovules différents.
Transferts nucléaires et clonages
D’autres expériences permettent de changer le patrimoine génétique des œufs. Le plus radical de ces changements consiste à détruire le noyau d’un œuf fécondé et à le remplacer par un noyau venant d’un autre embryon.
Les méthodes de transferts nucléaires ont été mises au point sur des oeufs beaucoup plus gros que ceux des Mammifères, les oeufs d’Amphibiens. Elles ont permis des expériences de clonage chez les crapauds, les Xénopes.
Un clone est une population (animale, végétale ou bactérienne) dont tous les individus sont génétiquement identiques : c’est un ensemble de sosies parfaits.
Pour réaliser une expérience de clonage, on part d’un embryon déjà formé de plusieurs cellules dont on isole et recueille tous les noyaux. Ces noyaux sont les descendants du noyau initial de l’œuf fécondé et sont donc tous génétiquement identiques entre eux. D’autres part, on prépare, une série d’œufs fécondés dont on détruit les noyaux. Ensuite, on injecte un noyau de l’embryon donneur dans chacun de ces oeufs.
Chez les Amphibiens, dans les expériences les mieux réussies, un certain pourcentage de ces oeufs se développe et produit donc autant de petits têtards tous rigoureusement identiques.
Les essais de clonage chez les Mammifères se sont heurtés à des difficultés techniques considérables. Ils ont toujours échoué chez la souris, mais ils ont parfois réussi chez la brebis et la vache où de petits clones de 3 à 5 animaux ont pu être obtenus.
Animaux transgéniques
Un autre type d’expérience combine les méthodes de génie génétique et de l’embryologie expérimentale. Dans les essais dits de transfection, on tente de transférer un type de gène, préalablement isolé, dans le noyau d’un œuf de Mammifère. L’animal qui résulte d’une transfection réussie est un animal transgénique.
Dans un cas spectaculaire d’expérience de transfection réussie (Brinster et al., 1983), le gène humain de l’hormone de croissance a été intégré dans les chromosomes d’un oeuf de souris. L’embryon opéré, transféré dans une mère porteuse, a produit une souris normale mais géante, de la taille d’un rat. Le gigantisme des animaux obtenus est lié au fait que le gène transféré n’a pas obéi aux signaux de contrôle normaux de son expression. Les animaux sont changés dans leur patrimoine génétique. Leurs descendants présenteront les caractéristiques de gigantisme induites par l’expérimentateur. Une nouvelle variété est créée.
Les méthodes de transfection se sont multipliées et raffinées ces dernières années. Au lieu d’utiliser une micropipette pour injecter un type de gène de manière massive et aveugle, on peut se servir d’autres vecteurs : rétrovirus, spermatozoïdes, cellules embryonnaires cultivées, préalablement transformés par l’incorporation du gène que l’on veut transfecter dans l’œuf. Ces méthodes permettent de n’insérer qu’une seule copie du gène dans l’embryon récepteur, de manière stable et parfois à l’endroit voulu. A cet égard, la découverte du virus AAV paraît prometteuse : il est inoffensif et s’insère toujours dans une certaine zone silencieuse, non codante du génome.
Enfin, la transgénose ouvre des perspectives biotechnologiques nouvelles. Par exemple, on tente d’obtenir des vaches transgéniques susceptibles de produire dans leur lait des protéines humaines à intérêt thérapeutique.
La différenciation cellulaire est le phénomène au cours duquel un type cellulaire est progressivement transformé en un autre type plus spécialisé.
1. Morphogenèse et homéostasie
Une cellule capable de se différencier, c’est à-dire de présenter des structures et des fonctions nouvelles, est en fait capable d’opérer des biosynthèses différentes d’une époque à l’autre de son existence. En particulier, elle synthétise des protéines nouvelles au cours des étapes de la détermination et de la différenciation cellulaires. L’expression de son génome varie donc avec le temps, suivant des séquences définies.
Ainsi, chez l’Homme, la durée de vie d’un érythrocyte (E) est d’environ 4 mois. Les cellules mères des érythrocytes (CME), situées dans les organes hématopoïétiques (rate, moelle osseuse), vont continuellement se diviser et se différencier de manière à compenser la perte des érythrocytes morts, le rapport CME/E étant constant chez les organismes sains. Pour tous les tissus susceptibles de se renouveler, on trouve ainsi des proportions définies et constantes de cellules différenciées (qui vont mourir) et de cellules non différenciées (qui vont remplacer les cellules mortes), ce qui entraîne la constance, ou homéostasie, du volume des tissus fonctionnels.
Qu’il s’agisse d’ailleurs d’homéostasie, d’embryogenèse ou de régénération, les cellules se différencient toujours de façon coordonnée, en harmonie avec l’organisme tout entier. Cette coordination implique l’existence de signaux circulant entre les cellules ; des mécanismes inter- et intracellulaires doivent permettre de répondre à ces signaux. Le manque de coordination conduit à des états pathologiques graves comme les malformations embryonnaires ou l’apparition de tumeurs.
La capacité de différenciation d’une cellule paraît tout à la fois dépendre :
a. de son information génétique, donc de son noyau ;
b. de la nature de son cytoplasme et de ses récepteurs membranaires ;
c. des signaux présents dans le milieu où elle vit ;
d. de la position qu’elle occupe dans l’organisme.
Le destin de chaque cellule dépend donc à la fois de ses propriétés intrinsèques et de son environnement.
2. Importance du noyau dans la différenciation cellulaire
De nombreux faits montrent l’importance du noyau dans le phénomène de la différenciation cellulaire. Nous citerons quelques exemples significatifs.
Les Acétabulaires sont des algues unicellulaires géantes où l’importance du noyau dans la différenciation a pu être démontré de manière directe. En effet, les différentes espèces d’Acétabulaires sont caractérisées par une ombelle sporifère, de forme spécifique et qui n’apparaît qu’à la fin de la croissance cellulaire. L’algue mesure alors environ 5 cm et le noyau est localisé dans le rhizoïde de fixation de la cellule. En coupant ce rhizoïde, on obtient une algue sans noyau. On peut assez facilement greffer le rhizoïde d’une espèce à une algue énucléée d’une autre espèce. Si l’on procède à ce type de greffe croisée entre de jeunes Acetabularia mediterranea et A. crenulata, les ombelles produites auront toujours la structure correspondant à l’espèce du noyau présent dans le cytoplasme hybride de la chimère obtenue.

Nombre de faits montrent l’importance des chromosomes dans l’embryogenèse. Citons en particulier :
l’hérédité des caractères paternels alors que seul le noyau du spermatozoïde pénètre dans l’œuf lors de la fécondation ;
l’effet létal des fécondations interspécifiques ;
les effets désastreux de l’aneuploïdie au cours de l’embryogenèse.
3. Importance du cytoplasme dans la différenciation cellulaire
Ici encore, des faits embryologiques nombreux témoignent en faveur de cette importance. En particulier, le fait que la détermination de régions cytoplasmiques existe dès le stade œuf fécondé indivis chez de multiples espèces. Des territoires purement cytoplasmiques, isolés, sont parfois capables d’effectuer des synthèses protéiniques sans présenter une réelle différenciation, les oeufs d’Oursins, capables de segmentation en l’absence de noyau.
Chez les Acétabulaires, le cytoplasme est capable d’accomplir une différenciation réelle, en l’absence complète de noyau. En effet, lorsqu’on sectionne l’ombelle d’une algue, la «plantule» régénère une nouvelle ombelle, normalement constituée. Lorsqu’on sectionne à la fois l’ombelle et le rhizoïde contenant le noyau, l’algue mutilée régénère une ombelle tout à fait normale. Les synthèses de protéines nécessaires à ce processus morphogène complexe peuvent donc se produire en l’absence du noyau et de l’information génétique qu’il contient. Ce qui n’empêche pas que cette information cytoplasmique soit supplantée par l’information nucléaire en cas de greffes croisées.
Les processus moléculaires sous-jacents aux capacités de différenciation autonome du cytoplasme sont encore fort mal connus. L’hypothèse la plus fréquemment invoquée est la présence dans ce cytoplasme de RNA messagers spécifiques, synthétisés dans le noyau mais capable de demeurer dans le cytoplasme pendant des temps longs, de l’ordre de quelques jours (cellules animales) à quelques semaine (Acetabularia).
Il est certain que la nature des gènes, la constitution génétique, conditionne l’activité cytoplasmique. Mais le cytoplasme contient des facteurs susceptibles d’agir sur l’activité du génome : l’expression des gènes est contrôlée par des facteurs cytoplasmiques. Ces facteurs sont certainement prédominants, sinon exclusifs, pendant la phase de segmentation de la morphogenèse.
L’expression modulée des gènes pourrait entre autres se traduire par l’acquisition de récepteurs protéiniques nouveaux au niveau de la membrane plasmique. La cellule deviendrait ainsi sensible à une nouvelle gamme de signaux extérieurs qui à leur tour pourraient moduler l’expression du génome.

















/https%3A%2F%2Fassets.over-blog.com%2Ft%2Fcedistic%2Fcamera.png)
/https%3A%2F%2Fprofilepics.canalblog.com%2Fprofilepics%2F2%2F2%2F220176.jpg)